
Haplogrupe mitohondrijske DNK čovjeka

Afrika, kolijevka početka migracija, je gore lijevo a Južna Amerike na krajnjoj desnoj strani.
Modeli migracije se zasnivaju na studijama mitohondrijske (matrilinijske) DNK.
Slova predstavljaju haplogrupe, a boje i brojevi predstavljaju hiljade godina prije sadašnjosti

Haplogrupe mitohondrijske DNK čovjeka, u genetici čovjeka, su haplogrupe koje su određene razlikama u mitohondrijskoj DNK. Haplogroupe su posebnp primjenljive u rekonstrukciji glavnih grana filogenetičkog stabla. Razumijevanje evolucijskih puteva Understanding the evolutionary path of the fem ženske linije bitno pomaže populacijskim genetičarima da trasiraju majčinsku liniju recentnog čovjeka do afričke i njegovog postepenog širenje širom Planete.
Oznake imena haplogrupa uopće, ne samo mitohondrijskih, idu od A do Z. Pošto su označene po redoslijedu otkrića, one (u smislu abecednog redoslijeda) ne odražavaju stvarne srodničke veze i odnose.
Hipotetska žena u korijenu svih tih grupa (znači samo mitohondrijskih DNK haplogroupa) je matrrilinijska najbliži pradavni zajednički predak danas živućih ljudi. Ona se obično naziva mitohondrijska Eva.
Evolucijski odnosi[uredi | uredi izvor]
Loze mitohondrijskih haplogrupa[uredi | uredi izvor]
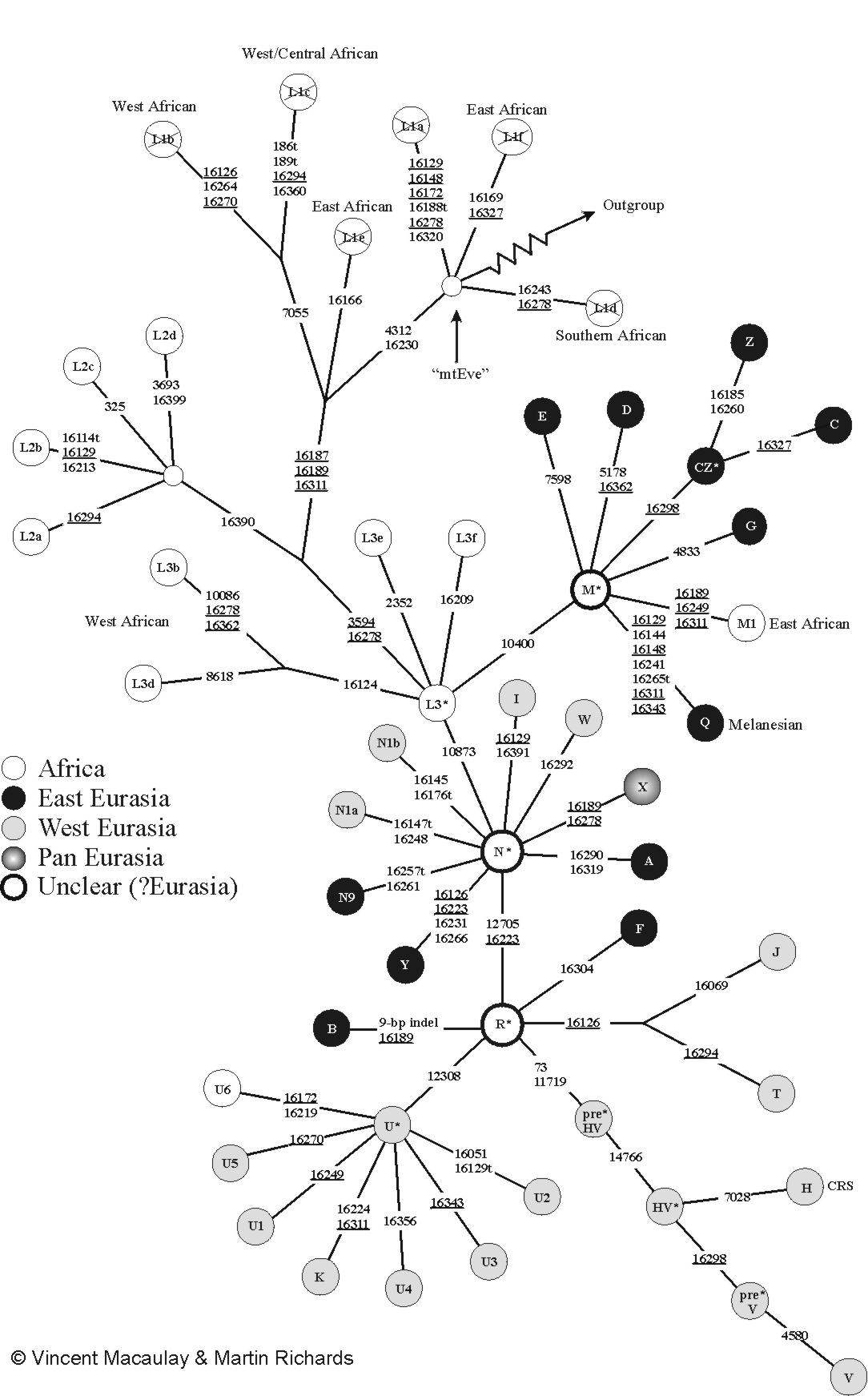
Filogenetičko stablo koje slijedi je bazirano na Ovenovom stablu iz 2009.[1] i naknadnim postepenim istraživanjima.
Hronologija razvoja haplogrupa[uredi | uredi izvor]
Evropske haplogrupe[uredi | uredi izvor]
Bryan Sykes tvrdi da recentni Evropljani potiču od sedam glavnih mitohondrijskih lozaza ali ostali sada pretpostavljaju da ih je 10-12. Ove dodatne "kćeri" općenito uključuju haplogroupe I, M i W. Nedavni radovi re-mapirali su evropske haplogrupe kao: H, J, K, N1, T, U4, U5, V, X i W.[2] Međutim, treba napomenuti da su haplogroupe proizvoljne kategorije definirane mutacijama, koje su i same rezultat brojnih slučajnih procesa. Moglo bi se reći da je razumno da postoji gotovo neograničen broj haplogroupa koje su prisutne u određenoj regiji. Osim toga, pripisujući specifično geografsko porijekla bilo kojoj haplogrupi, kao što su one koje su predložene u tabeli ispod, smatra se da je visoko spekulativno i da mogu biti 'pričanje priča' i izvan domena nauke. Isto se može reći da su rezultat prijedloga bliskih viđenja populacijskih genetičara.[3]
| Haplogrupa | Moguće vrijema nastanka (prije n hiljada godina) |
Moguće mjesto porijekla | Najviše frekvencije |
| N | 75.000 | Zapadna Azija, Indija ili Jučna Azija | |
| R | 70.000 | Zapadna Azija, Indija ili Jučna Azija | |
| U | 60.000 | Sjeveroistočna Afrika ili Jugozapadna Azija | |
| pre-JT | 55.000 | Srednji Istok | |
| JT | 50.000 | Srednji Istok | |
| U5 | 50.000 | Zapadna Azija | |
| U6 | 50.000 | Sjeverna Afrika | |
| U8 | 50.000 | Zapadna Azija | |
| pre-HV | 50.000 | Bliski Istok | |
| J | 45.000 | Bliski Istok ili Kavkaz | |
| HV | 40.000 | Bliski Istok | |
| H | preko 35.000 | Zapadna Azija | |
| X | preko 30.000 | Sjeveroistočna Evropa | |
| U5a1 | 30.000 | Evropa | |
| I | 30.000 | Kavkaz ili Sjeveroistočna Evropa | |
| J1a | 27.000 | Bliski Istok | |
| W | 25.000 | Sjeveroistočna Evropa ili Sjeveroistočna Azija | |
| U4 | 25.000 | Centralna Azija | |
| J1b | 23.000 | Bliski Istok | |
| T | 17.000 | Mezopotamija | |
| K | 16.000 | Bliski Istok | |
| V | 15.000 | Pirinejsko poluostrvo i širenje prema Skandinaviji | |
| H1b | 13.000 | Evropa | |
| K1 | 12.000 | Bliski Istok | |
| H3 | 10.000 | Zapadna Evropa (Španija) |
Afrička regija i mtDNK haplotipovi[uredi | uredi izvor]
Prosječno evolucijsko vrijeme datiranja afričke mtDNK varijacije procjenjeno je na 125.500-165.000 godina (Chen et al. 2000). Ove vrijednosti znatno premašuju dob svih azijskih i evropskih specifičnih mtDNK varijanti. Karakteristična makrohaplogrupa afričke mtDNK filogenije je haplogrupa L, čija je geografska distribucija uglavnom ograničena u istočnoj Africi i južnim dijelovima subsaharskog područja, a podijeljena je na L0-L6. Neke subhaplogrupe L specifične su za istočnu Afriku (npr. L0a, L0f, L5, L3g), neke za pojedine etničke skupine, kao što su L0d i L0k, pronađene isključivo među populacijama južne Afrike. Akumulacijom regionalnih varijacija prije 70-100.000 godina, iz drevne L0 haplogrupe rezultirale su u centralnoj i istočnoj Africi dvije varijante L2 i L3, čija je subsaharska disperzija (prije oko 60.000 godina) uglavnom nadmašila distribuciju LO haplogrupe. Afričku makrohaplogrupu L karakteriše osnovni ne-CRS plimorfizam na 3594 bp (tranzicija C→T) i polimorfizam +3592HpaI. Polimorfizmi +3592HpaI, tranzicija A→G na 10394 bp i +10394DelI karkaterišu sve mtDNK haplotipove koji pripadaju afričkoj-specifičnoj makrohaplogrupi L.
Jedna od najuticajnijih populacionih ekspanzija u posljednje 3-4.000 godina, koja se najvjerovatnije odrazila na reorganiziranje geografske substrukture afričke populacije, bila je ekspanzija Bantu populacija. U prilog toj činjenici je starosna dob glavnih tipova mtDNK osnivača zapadnog i istočnog afričkog podrijekla, (L02,L2, L3E i L5) koja je u skladu sa vremenom disperzije Bantu populacija iz zapadne, centralne i istočne Afrike prema jugu (Bandelt et al. 2001; Chen et al. 1995, Salas et al. 2002).
Afrički mtDNK genski fond je obogaćen i ne-afričkim mtDNK varijacijama (sl.10) Vrlo značajan uticaj na genetičku strukturu populacija sjeverne i istočne Afrike, imale su populacione ekspanzije sa Bliskog Istoka, iz perioda Gornjeg paleolita, iz kojih proizlaze haplogrupe N i M i njihove subhaplogrupe (Richards et al. 2003; Kivisild et al. 2004), zatim loze iz zapadne Euroazije u toku i nakon ledenog doba, post-glacijalne mtDNK haplogrupe iz francuskog-Cantabrian refugijuma i euroazijske linije unesene povratnim neolitskim ili skorašnjim migracijama (Rosa and Brehem 2011).
U Evropi, u ukupnom bazenu mtDNK varijacija loze afričke makrohaplogrupe L zastupljene su manje od 1% i ne pokazuju signale da su evulirale na evropskom kontinetu. Većina ovih loza (oko 65%) potiče iz skorijeg vremenskog perioda, perioda romanizacije, arapskog osvajanja Iberijskog poluotoka i Sicilije, dok ostale loze formiraju evropske specifične subhaplogrupe, otkrivajući da se protok gena iz subsaharske Afrike prema Evropi desio prije oko 11.000 godina (Cerezo et al. 2012).
Haplogrupa koja se izdvaja iz spektra afričkih mtDNK haplogrupa i smatra izvornom nukleotidnom mitohondrijalnom sekvencom, zajedničkim MRCA svih ne-afričkih mtDNK loza je L3. Ovu afričku haplogrupu karakteriše polimorfizmom -3952HpaI (Chen et al. 1995). MtDNK haplotipovi koji imaju ovaj polimorfizam odgovorni su za razdvajanje modernih sub-saharsko afričkih populacija od azijskih i evropskih.
Divergencijom afričke haplogrupe L3 prije 70-100.000 godina rezultirale su se dvije super-haplogrupe M i N, predstavnici mitohondrijalnih varijacija izvan Afrike (isključujući potomke migracija iz Afrike u posljednjih nekoliko hiljada godina). Ove loze emigrirala su iz Afrike prije oko 60-65.000 godina prema Euroaziji i Novom svijetu (Quintana-Murci et al. 1999; Mishmar et al. 2003; Macaulay et al 2005). Od inicijalnih M i N makrohaplogrupa u pojedinim regijama euroazijskog kontinenta nastale su nove, danas regionalno-specifične subhaplogrupe.
Makrohaplogrupa M[uredi | uredi izvor]
Makrohaplogrupa M predstavlja ancestralnu haplogrupu istočno-euroazijskog porijekla. Prvobitno je definisana polimorfizmom +10394Ddel, (transverzija na A→G 10398) i +10397AluI (tranzicija C→T na 10400bp) (Wallace et al. 1995). Veliki diverzitet i prostorni raspored sugeriraju da se azijska varijanta haplogrupe M odvojila od istočnog-afričke haplogrupe M1 (varijanta haplogrupe M u Africi), prije više od 50.000 godina.
Oko 60% mtDNK autohtonih populacija istočne Euroazije pripadaju makrohaplogrupi M i njenim subhaplogrupama. Najćešće subhaplogrupe pripadajuće makrohaplogrupe M su C, Z, D, G, E i Z. Najfrekventnije haplogrupe sjeverne, centralne, istočne Azije, a također i Amerike su C i D. Noviji podaci ukazuje da su se ove mtDNA grane proširile prije posljednjeg glacijalnog maksimuma (engl. Last Glacial Maximum-LGM), sa svojim najstarijim lozama iz istočne Azije, a disperzija većine sjeverno-azijskih varijanti haplogrupa C i D se desila nakon LGM, ukazujući na naseljavanje ovog dijela Azije u post-glacijalnom periodu.
Južna Azija reprezentira sa oko trećinom specifičnih subhaplogrupa M. Najveći diverzitet makrohaplogrupe M zabilježen je u Indiji, gdje su prema dosadašnjim podacima identifikovane sljedeće subhaplogrupe: M2, M3, M4, M5, M6, M18, M25, M30-41, M48-50, označene kao indijsko-specifične loze. Drevna indijsko-specifična haplogrupa je M2, najstarija loza sa procjenom starosti oko 49.686 godina. MtDNK genofond južne Azije sadrži autohtone haplogrupe ukupnog mtDNK stabla proizašle direktno iz makrohaplogrupe M, kao N i R. Jedina indijsko-specifična subhaplogrupa koja ne proizilazi iz bazalne mitonodrijalne osnove je U2 koja sadrži tranziciju na 16051 bp u kontrolonom regionu, polimorfizam koji nije jedinstven za osnovu haplogrupu U. Vrijeme divergencije ove U2 je oko 50.000 godina.
U odnosu na najveći diverziteta i visoku prostornu distribuciju u Aziji, makrohaplogrupa M i njene subhaplogrupe u Evropi se tek sporadično pojavljuju (uočena frekvencija u zapadnoj i centralnoj Evropi je manja od 2%). Njeno prisustvou Evropi može biti uticaj vjerovatno skorijih historijskih ili individulanih migracija.
Za determinaciju strukture mtDNK loza prisutnih u Aziji korištene su brojne studije analiza kompletnih i parcijalnih sekvenci kodirajuće regije mtDNK. Kombinovanjem objavljenih podataka, sumirali su topologiju istočno, južno i zapadno-euroazijskih mtDNK haplogrupa uz ključne haplogrupe Australije i Nove Gvineje.[4]
Makrohaplogrupa N[uredi | uredi izvor]
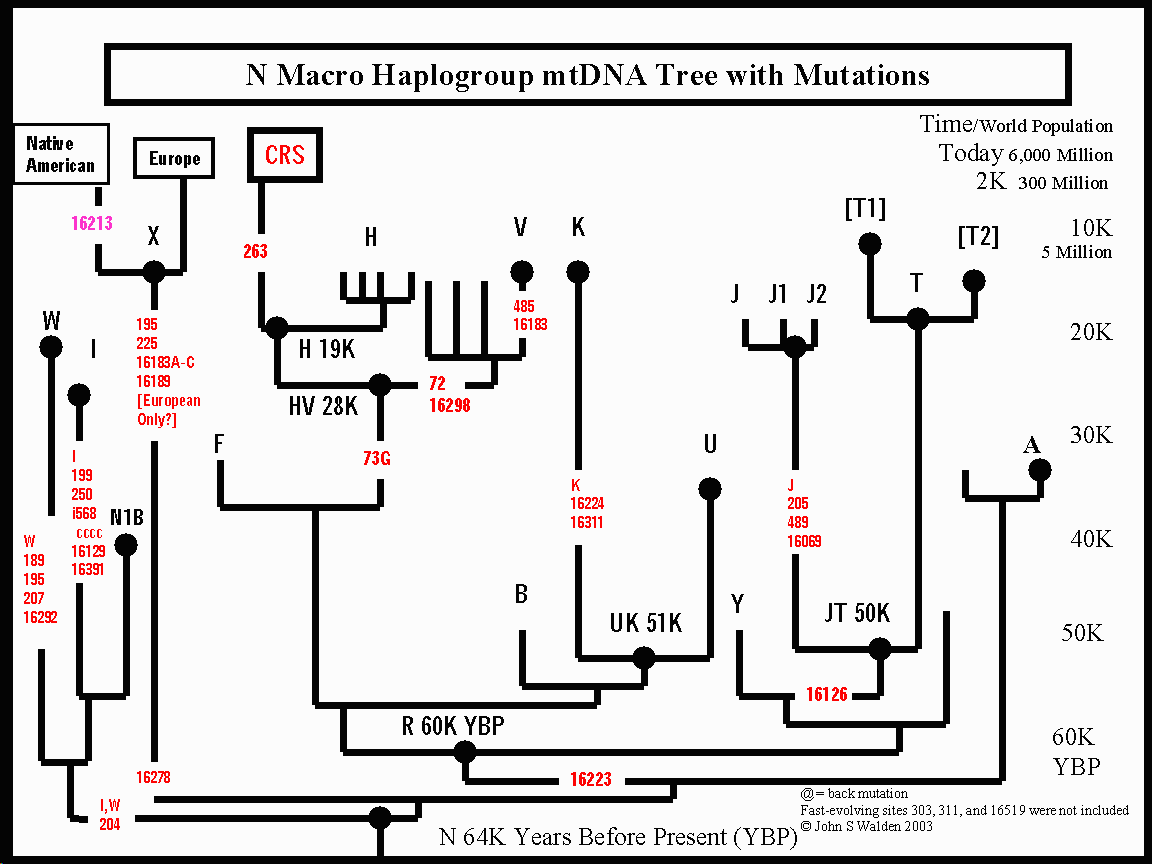
Makrohaplogrupa N obuhvata ostatak azijskih i evropskih varijacija, te pripada evroazijskom porijeklu. To znači da filogenetičko stablo makrohaplogrupe N obuhvata sve evroazijske haplogrupe koje karaketriše polimorfizam -10397AluI i +10394Ddel (Maji et al. 2008). Prisutna je u skoro svim ne-afričkim populacijama i prema današnjoj distribuciji je ne-afričkog porijekla. Subhaplogrupe haplogrupe N uočene u Africi su vjerovatno imigranti, pripadnici haplogrupe U (kao npr. U6, koja je dominantna u sjevernoj Africi).[4]
Filogeografija makrohaplogrupe N pokazuje specifičnu-geografsku distribuciju, obuhvatajući istočno (A, B, Y i R9) i zapadno (N1, N2, JT, HV, U, W) evroazijske varijante. Za zapadno-evroazijske populacije karakteristična je uniformna distribucija svih šest osnovnih subclada N1, N2, X, pre-HV, JT i U izvedenih iz glavnih izvornih haplogrupa N i R, a u koje se svrstavaju praktično sve evropske haplogrupe: H, I, J, K, R, T, U, V W i X, čija je distribucija uglavnom ograničena na zapadu Euroaziju i sjevernu Afriku (osim U, koja je pronađena i u Indiji). U Indiji većina subhaplogrupa N su varijante R uključujući R5, R6 i U2. R5, U2a, U2b i U2c, i čine oko 15% mtDNK varijacija južne Azije, čije je koalescentno vrijeme procjenjeno preko 50.000 godina. Ove haplogrupe predstavljaju drevne indijsko-specifične haplogrupe izvedene iz makrohaplogrupe N, dok su nove indijsko-specifične zasnovane na subhaplogrupama N5,R7, R8, R30, i R31. Također, specifične subhaplogrupe osnovnih M i N makrohaplogrupa karakteriziraju i populacije Nove Gvineje i Australije.
Evropska regija i zapadno-evroazijske varijante mtDNK[uredi | uredi izvor]
Molekulske analize ukazuju da se najviše varijacija tokom evolucije evropske mtDNK akumuliralo u sekvencama postepenim širenjem loza iz seta osnivačkih mtDNK makrohaplogrupa (N i R) u toku i poslije procesa kolonizacije evropskog kontinenta. Većina evropskih mtDNK varijacija pripada osnovnim haplogrupama pre-HV, JT i U i njihovim subhaplogrupama, proizašlim iz makrohaplogrupe R, a ostali manji dio pripada minornim subhaplogrupama N1, N2 i X. Oko 80-90% evropskih mtDNK varijacija klasificirano je u jednu od devet H, I, J, K, T, U, V, W ili X, koje su označene kao zapadno-euroazijske haplogrupe. Prema jednoj od prvih analiza (Torroni et al. 1996) koja se bavila determinacijom geografskih mtDNK varijanti, šest haplogrupa H, I, J, K, T, W predstvaljene su kao esencijalne evropske haplogrupe.[5] Osim navedenih glavnih zapadno-euroazijskih haplogrupa u evropskim populacijama je zastupljeno i nekoliko reprezentativnih azijskih (M) i afričkih (L1 i L2) varijanti, čije je porijeklo na evropskom tlu vjerovatno posljedica nekih tipova sporadičnih individualnih disperzija.
Prema dosadašnjim istraživanjima evropskih mtDNK varijacija, skoro svi evropljani dijele isti skup mtDNK haplogrupa kao i populacije B. Istoka, a koje su obično odsutne u subsaharsko-afričkim i istočno-afričkim populacijama. Kompletna mtDNK filogenija i geografska distribucija ukazuju da su evropske varijacije osiromašene u broju osnivačkih bazalnih mtDNK loza u odnosu na mtDNK varijacije B. Istoka. Smanjena zastupljenost bazalnih mtDNK varijacija u ukupnom evropskom mitohondrijalnom genofondu vjerovatno je rezultat periferne uloge pionirskih migracija, koje se u Evropi pojavljuju tek poslije naseljavanja B. Istoka. Vrijeme datiranja postojećih bazalnih (osnivačkih) mtDNK varijacija u evropskim haplogrupama je procjenjeno na oko 40-50.000 godina (Dansgaard et al. 1993). Prema literaturnim podacima još uvijek nije precizno jasno u kojoj mjeri i na koji način su prve migracije modernih ljudi iz Afrike oblikovale mtDNK bazen modernih evropljana i na koji način su kasnije migracije iz B. Istoka zamijenile potomke prvih evropskih naseljenika.
Osim haplogrupa V i U5, za koje se predpostvalja da su autohtonog evropskog porijekla, sve ostale evropske haplogrupe unesene su iz B. Istoka protokolonizacijom prije oko 45–50.000 godina. Dalja divergencija mtDNK haplogrupa u Evropi rezultirala je najviše tokom migracija u periodu postglacijalnih ekspanzija (prije oko 14.000 godina) i neolitskih disperzija prije oko 10.000 godina iz Bliskog Istoka.
U popularnoj knjizi B. Sykes (2001) ,,The Seven Daughters of Eve,, evropske mtDNK haplogrupe, popularno su nazvane: Helena (H), Velda (V), Jasmine (J), Tara (T), Xenia (X), Katrine (K), Ursula (U).[4]
Haplogrupa V[uredi | uredi izvor]
Haplogrupa V predstavlja autohtonu evropsku mtDNK haplogrupu. Samo oko 4% današnjih modernih evropljana su nosioci ove haplogrupe. Prisustvo trancizije (T→C) na nukleotidnoj poziciji 16298 HVRI prvi put je uočeno u studiji Torroni et al. (2001), u populacijama zapadne Evroazije i sjeverne Afrike, koja je predhodno predstavljala marker za determinaciju ove haplogrupe. Sve mtDNK loze koje posjeduju ovaj polimorfizam mogu pripadati jednoj od dvije haplogrupe, ili haplogrupi V ili filogenetski starijoj haplogrupi pre-V. Geografska distribucija i evolutivno vrijeme haplogrupe pre-V sugerišu o evropskom porijeklu prije LGM (engl. Last Glacial Maximum), pokazujući tedenciju širenja ostatkom Evrope i sjeverozapadne Afrike. Procjenjeno prosječno evolutivno vrijeme haplogrupe V (derivata pre-V haplogrupe) za populacije jugozapadne Evrope je 16.300(+/-4.800) godina, ukazujući na njeno pojavljivanje ubrzo nakon LGM, dok za istočne populacije ono iznosi 8.500(+/-2.300), što je vjerovatano u asocijaciji sa postneolitskim ekspanzijama iz zapadne Evrope prema istočno/centralnoj Evropi. Prosječan broj nukleotidnih razlika u sekvenci HVRI, sugeriše da je Iberijsko popuostrvo/južna Francuska, mjesto porijekla autohtone evropske haplogrupe V, iz kojeg se dalje širila u postglacijalnom periodu ostatkom Evrope.
Haplogrupa V pokazuje dva vrha frekvencije, jedan u jugozapadnoj Europi, a drugi u sjevernoj Skandinaviji. Visoka frekvencija (10-12%) zabilježena u populaciji Baski u Iberijskom populostrvu (jugozapadna fEvropa), i vjerovatno je rezultat genetičkog drifta ili njihovog miješanja sa kasnijim migrantima nosiocima haplogrupe V. Veliki procenat haplogrupe V (oko 40%), zabilježen je u Sammi populaciji (sjeverna Skandinavija), zatim među Berberima sjeverne Afrike (8-11%). Ova haplogrupa je jedna od najbolje okarakterisanih evropskih loza mitohondrijalnog genskog toka. Iako samo oko 4% današnjih modernih ljudi evropskog porijekla pripada haplogrupi V, novije publikacije, izvještavaju o porastu ograničene distribucije haplogrupe V modernim evropskim popualcijama, koja je reflektovana obrascima postgalcijalne kolonizacije iz Iberijskog poluostrva. S obzirom, da povećava gradijent svoje frekvencije od zapada prema ostatku Evrope predstavlja marker za procjenu pravca kretanja populacija iz zapadnih okruženja u vremenu postglacijalne re-kolonizacije Evrope.
Haplogrupa U[uredi | uredi izvor]
Jedna od prvih filogenetičkih analiza varijacija mtDNK u populacijama zapadne Evrope, sugerirala je, da se najstarija evropska mitohondrijalna haplogrupa U (sa procjenom starosti i preko 50.000 godina), pojavila sa najranijijim migracija iz B. Istok. Prema dosadašnjim studijama, ova haplogrupa je reprezentovna brojnim subhaplogrupama, od kojih je svaka okarakterisana zasebnim nukleotidnim razlikama, geografijom i historijom. Npr. subhaplogrupe U1-U5 su pronađene u zapadno-evropskim populacijama, subhaplogrupa U6 je karaketristična za afričke populacije, a U7 je tipična za populacije Jordana , Saudisjke Arabije i Irana.
U subklasterskoj diversifikaciji haplogrupe U, poziciju najstarije evropske subhaplogrupe zauzima U5 (45-50.000 godina) (Richards et al. 1996;1998; 2000), koja se vjerovatno širila iz B. Istoka prvim evropskim naseljenicima u ranom gornjem paleolitu (engl. Early Upper Paleolithic), zajedno sa Aurignacian kulturom. Ovoj haplogrupi pripada i važna uloga u postglacijalnoj ekspanziji populacija iz jugozapadne Evrope.
Značajnu ulogu u genetičkoj strukturi evropskih populacija ima i haplogrupa U4, determinanta srednjeg gornjeg paleolita (engl. Middle Upper Paleolithic-oko 26000). Evolutivno vrijeme procjenjeno na osnovu divergencije mtDNK tipova u B. Istoku i Evropi je oko 25.650-20.400, sugerišući da je B. Istok mjesto porijekla ove subhaplogrupe (Richards et al. 2000). Prema podacima geografske distribucije u evropskim populacijama najveća učestalost (2- 20%) je u istočnoj Evropi (u populacijama Volga–Ural i Cis-Ural regiona) (Bermisheva et al. 2002). Osim evropskih populacija, ova haplogrupa je prisutna u relativno velikoj frekveciji među autohtonim populacijama sjeveroistočnog Sibira, Mansi, Nganasans i Kets (16-29%) (Debrenova 2002a, 2002b). Rekonstrukcija filogenetskih relacija između U4 mtDNK tipova u današnjim populacijama istočne Evrope, Urala i sjeverozapadnog Sibira ukazuje da je divergencija ovih subhaplogrupama u istočnoj Evropi počela u gornjem paleolitu (tačnije prije oko 23.000 godina).
Značajna determinantna mtDNK toka u Evropi je i subhaplogrupa komponenta srednjeg gornjeg paleolita. . Ova haplogrupa je rijetko prisutna u evropskim populacija, ali je jedna od dominantnijih haplogrupa u Indiji. Zapadno-evroazijska forma klastera U2 uključuje transverziju na nukleotidnoj pozici 16129, koja je odsutna u Indiji. Još jedna komponenta srednjeg gornjeg paleolita značajna za genetičku strukturu evropskih populacija je U1a grana U1 haplogrupe. U svom radu tragajući za evropskim osnivačkim linijama mtDNK toka u B. Istoku Richards et.al. (2000) prisustvo haplogrupe U1 u Evropi povezuje sa ekspanzijama iz srednjeg gornjeg paleolitika prije oko 26.000 godina.
Haplogrupa H[uredi | uredi izvor]
Danas je oko 30-50% evropskih mtDNK linija (izuzimajući Sammi populacije) klasificirano kao haplogrupa H. Najfrekventnija je u zapadnoj i sjevernoj Evropi (40-50%), dostižući maximum u Baski populacijama (oko 60%), a srednje frekventna je u južnom i jugozapadnom dijelu kontinenta (20%-40%). Iako je specifična haplogrupa evropskog kontinenta, ova haplogrupa pokazuje i relativno široku geografsku distribuciju, pojavljuje se i u B. Istoku (oko 25-30%) i Kavkazu, sjevernim dijelovima Indije, centralne Aziji, ali u dosta manjoj frekvenciji (5-10%).
Haplogrupa H je porijeklom iz srednjeg Istoka (prije oko 25-30.000 godina), u Evropi se pojavljuje i širi u asocijaciji sa drugim valom paleolitskih migracija, vjerovatno sa difuzijom Gravettian tehnologije, što ukazuje da je bila uključena u post-glacijalnu ekspanziju humanih populacija iz ledenih refugija poslije LGM. Jedna od najpriznatijih studija sugerira da je haplogrupa H glavna komponenta jedne od četiri prahistorijske migracije iz B. Istoka prema Evropi iz perioda kasnog gornjrg paleolita, (prije oko 14.000). Koalescentno vrijeme većine subhaplogrupa H korespedentno je pojavljivanju njihove ekspanzije u postglacijalnom periodu, te haplogrupa H predstvalja jednu od glavnih determinanti rekolonizaciji Evrope.Molekularna disekcija haplogrupe H sugeriše na pojavljivanje subhaplogrupa mlađe dobi, koje doprinose dosta preciznijoj spoznaji o prahistorijskim i historijskim događajima na prostoru Evrope. Najfrekventnija je sub-haplogrupa H6 u zapadnoj Evroaziji, subhaplogrupa koja je bila najznačajna u dosadašnjim studijama uočenih varijacija mt genoma u Evropi. Prvo kompletno sekvencioniranje 62 mtDNK uzorka, uz primjenu RFLP markera −7025AluI i −14766MseI opisano je 2001. Prema trenutnoj haplogrupnoj nomenklaturi Cambridge Reference sequence (CRS) pripada haplogrupi H, odnosno subhaplogrupi H2B.[4]
Haplogrupa I[uredi | uredi izvor]
Haplogrupa I je pronađena širom Evrope, ali u dosta ograničenoj frekvenciji (oko 2%). Prema rezultatima novije studije Oliveri et al. (2013) hapalogrupa I potiče iz B. Istoka iz perioda ,,LGM,, odakle se dalje širi u Evropu postgalcijalnom ekspanzijom. Realtivno visoka frekvencija u Evropi uočena je u istočnom Mediteranu (4,7%), a zabilježena prosječna frekvencija u evropskim populacijama je oko 1%.
Haplogrupa N1a[uredi | uredi izvor]
Prema filogenetičkim podacima prvi evropski farmeri su najvjerovatnije bili nosioci subhaplogrupe N1a makrohaplogrupe N. Subhaplogrupa N1a predstavlja važan genetički marker za procjenu prisustva ranih neolitskih farmera u Evropi. Evropskim kontinentom širila se sa Linearbandkeramik (LBK) kulturom, koja se vezuje za prve poljoprivredne zajednice u Evropi prije 7.500 godina (Hakk et al. 2005). Nekada bila dominirajuća haplogrupa među neolitskim farmerima u centralnoj Evropi, zauzimajući oko 25% genskog toka (Palanichamy et. al. 2010). Cjelokupna filogeografska distribucija N1a subklastera i njihovo koalescentno vrijeme 19.600-23.500 sugeriše da se inicijalna diversifikacija N1a pojavila u B. Istoku, nakon čega je uslijedila ka zapadu disperzija predačkih pojedinačnih subhaplogrupa u južnoj Evropi i preko stepskih zona Centralne Azije ka centralnoj Evropi. N1a tip sadrži tri filogeografska subklastera: istočno afričko/južno azijski, evropski i centralno-azijski. Evropske i centralno-azijske varijante imaju karakteristični ,,motif,, 16147A u kontrolnom regionu, dok afričko-azijsku varijantu karakteriše mutacija na 16147G.
Na osnovu N1a haplogrupne filogenije i filogeografskih informacija, rezultati novijih studija su nešto drugačiji u odnosu na hipotezu da su N1a linije bile donesene u centralnu Evropu neolitskim farmerima iz B. Istoka demskom difuzijom. Filogeografska analiza N1a linija (Palanichamy et.al. 2010) naglašava da N1a linije evropskih farmera mogu imati različito porijeklo, iz različitih izvora – iz istočne Evrope, da Bliskog Istoka preko Južne Evrope (za N1a1b i vjerovatno N1a1a3) i iz lokalnog centralnog evropskog izvora (za N1a1a2). Ovi podaci ukazuju da je neolitska tranzicija dosta kompleksan proces.
Haplogrupa J[uredi | uredi izvor]
Haplogrupa J bila je prvi put opisana u jednoj studiji, u kojoj je okarakterisana polimorfnim pozicijama -13704BstNI i 16065HinfI (gubitak restrikcionog mjesta na nukelotidnim pozicijama 13704 i 16065). ). Izvorna haplogrupa J vodi porijeklo iz Bliskog Istoka (prije oko 50.000 godina), a u Evropu je donesena neolitskim migracijama. U prilog ovoj činjenici ide i procjenjen nešto veći diverzitet haplogrupe J u Bliskom Istoku (0,987), u odnosu u Evropi (0,899). Danas je pretežno prisutna u Evropi. Jedna činjenica koja razlikuje haplogrupu J od ostalih evropskih haplogrupa je tranzicija A>G na 73bp, koja nije specifična za haplogrupu J, kao za ostale evropske-specifične haplogrupe (Serk 2004). U evropskim populacijama prosječna učestalost je oko 10%, dostižući vrhunac u frekvenciji kod Španjolaca (i do 12 %) i kod Italijana (18 %). Prosječna frekvencija ove haplogrupe zabilježena je u području Bliskog Istoka (oko 12%), dostižući visoku vrijednost na Arapskom poluotoku među Beduinima (oko 25%).
MtDNK filogenija ove haplogrupe je dugo bila zasnovana na podacima parcijalnih analiza hipervarijabilnih regiona i RFLP podacima. Njena filogeografija je dosta dobro inetrpretirana kao glavni genetički argument za širenja populacija modelom ,,demic difuzije,, iz Anatolije i Levanta. Logan et al. (2010) godine predstavljaju filogeniju haplogrupe J na osnovu podataka 111 sekvencvi kompletnog mitohondrijalnog genoma. U novijim evolutivnim studijama pronađen je još veći broj specifičnih polimorfizama koji povećavaju filogenetsku rezoluciju topologije haplogrupe J.
Haplogrupa T[uredi | uredi izvor]
Haplogrupa T predstavlja jednu od tipičnih evropskih haplogrupa, koja se zajedno sa haplogrupom J širila neolitskom ekspanzijom iz Bliskog Istoka. Najprihvaćenija teorija je, da su ,,Tara,, ljudi došli sa Bliskog Istoka populacijama farmera. Najveća frekvencija ove haplogrupe je nađena u populacijama srednjeg Mediterana. Distribuirana je Evropom sa prosječnom frekvencijom oko 9% i pretežno koncentrisana u sjeverozapadnoj Italiji.
Haplogrupa K[uredi | uredi izvor]
Haplogrupa K predstavlja ogranak haplogrupe UK i jedna od šest specifičnih haplogrupa svojstvenih evropskim populacijama. Ova haplogrupa je uglavnom evroazijski haplotip i pretpostavlja se da se pojavila među prvim haplogrupama u procesu ekspanzije humanih populacija kroz Evropu, poslije posljednjeg glacijalnog maksimuma (LGM) (oko 15-20.000 godina) (Behar et al. 2006). Na osnovu procjenjene starosti sugeriše se da su individue sa ovom haplogrupom sudjelovale u pre-neolitskoj ekspanziji. Oko 32% haplotipova modernih ljudi aškenaskog jevrejskog porijekla su pripadnici haplogrupe K.[6]
Haplogrupa W[uredi | uredi izvor]
Karakteristična evropska haplogrupa W, prisutna je u Evropi sa veoma malom učestalošću (1-3%). Osim Evrope, pojavljuje se i u zapadnoj i južnoj Aziji. Rezultati novijih studija sugeriraju na njeno porijeklo iz B. Istoka iz perioda LGM. S obzirom na oskudicu podataka, budući radovi bi trebali što preciznije definisati historijsku distribuciju ove haplogrupe i preciznije odrediti njenu ulogu u procesu naseljavanja Evrope.
Haplogrupa X[uredi | uredi izvor]
Haplogrupa X, iako je distribuirana u skoro svim evropskim populacijama, zbog svoje niske učestalosti predstavlja mali udio u ukupnim evropskim mtDNK varijacijama. Relativno visoka učestalost ove haplogrupe u evropskim populacijama smatra se vrijednost od 4 do 5%, koja je zabilježena među populacijama istočnog i srednjeg Mediterana, te u Makedoniji. Najveća frekvencija ove haplogrupe je uočena u sjevernom Kavkazu, Istočnoj Evropi, Sibiru, i centralnoj Aziji. Haplogrupa X je također jedna pet haplogrupa pronađenih kod autohtonih Amerikanaca. Iako je zastupljena samo u 3% u autohtonim populacijama Amerike predstavlja glavnu haplogrupu sjeveroistočnog dijela sjeverne Amerike, gdje je među Algonquian populacija prisutna sa oko 25%. U nešto manjem procentu (11-15%) je prisutna u zapadnim i južnim dijelovima sjeverne Amerike.
Iz glavne haplogrupe N prije oko 20-30.000 godina divergirala je genetska sekvenca haplogrupe X dajući dvije subhaplogrupe, X1 i X2. Subgrupa X1 je manje frekventna i ograničena je na sjevernu i istočnu Afriku i Bliski Istok (Derenko et al. 2001). Sub-grupa X2 pojavljuje se u ekspanziji populacija poslije posljednjeg glacijalnog maksimuma u zapadnoj Evroaziji.[4]
Koncept mitohondrijske Eve[uredi | uredi izvor]
Prema dosadašnjim studijama i hipotezama, najpopularnije su dvije teorije o porijeklu i migracijama modernih ljudi: model Out Africa ili Single, recent origin model. Prvi model je ćešće prihvaćen od strane brojnih istraživača, zbog većeg broja genetičkih dokaza koji idu u prilog ovom modelu. Model tumači da se H. sapiens prvi put pojavio u Africi prije oko 100-200.000 godina, da mu je predhodio duži period nagomilavanja genetičkog diverziteta i da se širio ostatkom svijeta prije oko 100.000 godina. Prema ovoj hipotezi bilo je vrlo malo ili skoro nikako genetičkog miješanja sa arhaičnim formama roda Homo, a formiranje regionalnog-geografskog diverziteta među populacijama rezultat je selekcije i genetičkog drifta.
Model Multiregional continuity, (multiregionalni kontinuitet) sugeriše da porijeklo svih modernih humanih populacija seže u period kada se pojavio H. erectus prije oko 2 miliona godina (računajući i afričkog predstavnika H. ergaster). Prema ovoj hipotezi svi moderni ljudi su evoulirali istovremeno iz arhaičnih formi u različitim dijelovima svijeta, a kao posljedica pojavljivanja ranih populacija H. erectus-a pojavljuju se međupopulacijski diverzitet u različitim geografskim regionima i uporno se proteže skroz moderne populacije. Ovo ,,protezanje,, je označeno kao regionalni kontiniutet (engl. regional continuity). Koncept je zasnovan uglavnom na analizi fosilnih ostataka, koji izvještavaju o dokazima morfološkog i kuturnog kontinuiteta između arhaičnih formi i modernih ljudi izvan Afrike. U svojoj izvornoj interpretaciji multiregionalna hipoteza je pretpostavljala postojanje vrlo ograničenog genskog protoka između različitih geografskih populacija, međutim, od tada je modifikovana i tumači da je genski protok bio veoma važna komponenta evouliranja humanih populacija. Najnovija formulacija, sada je poznata kao hipoteza ,,multiregional evolution,,, koja ukazuje na balansirani genetski tok između geografskih odvojenih populacija i održava regionalni anatomski integritet.
Osim što je podržan na osnovu paleoantropoloških, zatim podataka specifičnih autosomalnih genetičkih markera, model Out Africa modernih humanih populacija, prihvaljiv je i sa aspekta analize mtDNK varijacija. Prve aplikacije podataka humanih mtDNK varijacija korištene za analiziranje tempa i modela evolucije primata, pojavljuju se početkom 1980-tih godina. Međutim, prvi put zvanično priznati podaci primjene mtDNK polimorfizama (zasnovani na restrikcijskom mapiranju) za kreiranje humanog filogenetskog stabla proizilaze 1987. godine i sugerišu koncept mitohondrijska Eva. Koncept je zasnovan na teoriji da svi moderni ljudi (po ženskoj liniji) vode porijeklo od jednog zajedničkog mitohondrijalnog pretka, popularno nazvan ,,mitohondrijlana Eva,, za koju se pretpostavlja da je živjela u Africi prije oko 150-200.000 godina, a čija se skupina danas smatra zajedničkim pretkom svih modernih ljudi po majčinskoj liniji (mtMRCA-mtMost Recent Common Ancestor -), a ne zajedničkim pretkom (MRCA-Most Recent Common Ancestor). Postavljeni koncept podržava model ,,out Africa,, sugerišući na prvo pojavljivanje H. sapiens (po ženskoj liniji) u Africi prije oko 150-200.000 godina. Također, brojne kasnije studije o mtDNK varijacijama podržale su hipotezu ,,out Africa model,, što je generalno prihvaćeno od strane brojnih istraživača. Inicijalna afrička populacija pripadala je ancestralnoj haplogrupi L koja se u Africi pojavljuje prije više od 150.000 godina (sl.17).
Novije analize procjene evolutivnog doba mtMRCA zasnovane na primjeni različitih metoda analize mtDNK varijacija (isključujući D petlju) procjenjuje da je vrijeme datiranja mMRCA oko 170.000 (+/- 50 ili 70) (Ingman et al. 2000). Procjena evolutivnog doba mtMRCA za mlađe klastere koje sadrže obje i afička i ne-afrička sekvence je oko 50,000 (+/- 27,500).
Vanafrički tokovi mitohondrijskih gena[uredi | uredi izvor]
Tačno vrijeme disperzije anatomskih modernih ljudi iz Afrike fundamentalno je pitanje u studijama humane evolucije. Najranije migracije iz Afrike vezuju se za prehistorijski period, još od vremena H. erectus-a prije oko 1,8 miliona godina. Analize brojnih klasičnih genetskih markera, kao i arheološki i paleoantropološki podaci ukazuju da je većina emigracija anatomskih modernih H. sapiens-a dovela do zamjene svih ranijih formi H. erectus-a kao i arhaičnih predstavnika (naendertalaca). Fosilni ostaci pokazuju da su se anatomski savremeni ljudi širili iz Afrike u pravcu B. Istoku prije oko 100-130.000, a nakon toga iz tih regija uslijedile su disperzije prema ostalim dijelovima svijeta. Podaci genetskih analiza recentnih populacija sugerišu na vremenski nešto kasnije migracije i različite rute disperzija. U nastavku ovog odjeljka ćemo pokušati predočiti dosadašnja saznanja o genskom toku humanih populacija iz Afrike i njihovoj disperziji prema ostalim regionima svijeta (sa posebnim osvrtom naseljavanja Evrope), iz perspektive mtDNK, a prema rezultatima novijih svjetskih istraživanja.
Rezultati brojnih analiza mtDNK sugerišu da su posljednje dvije disperzije iz Afrike (,,južna ruta,, prije oko 70.000 godina i ,,sjeverna ruta,, prije oko 50.000 godina) imale značajan uticaj na kompoziciju genetičke strukture svih ne-afričkih populacija. Također, mtDNK geneologija regiona Azije ukazuje na dvije velike migracije iz Afrike. Južna ruta ,,Route Southern Coastal,, (SCR) predstavlja disperzije iz Horn Afrike (Rog Afrike-istočna Afrika) duž obale Indijskog okeana (obala južne i jugoistočne Azije) prije oko 60-70.000 godina (Fernandes et al. 2012; Soares et al 2013; Torroni et al 2006. Ova ruta je obilježena ekspanzijom haplogrupe M (30-70.000 godina) koja se širila iz regiona istočne Afrike do Indije i jugoistočne Azije, a kasnije i do Australije (prije oko 50-60.0000) (sl.17.18) (Bowler et al 2003; Quintana-Murci et al 1999). Prema dosadašnjim studijama mtDNK varijacija genofond jugoistočne Azije ima ,,visoku,, bazalnu diversifikaciju makrohaplogrupe M, što ide u prilog hipotezi da je Azija vjerovatno bila prvi geografski prostor naseljen migracijama iz istočne Afrike i rapidnoj kolonizaciji duž obala južne Azije prije oko 60-70.000 godina. Također, ,,visoka,, bazalna diversifikacija sugeriše da Indijski subkontinent ima ,,zatvoreno,, porijeklo haplogrupe M i da je njeno pojavljivanje u Africi i B. Istoku rezultat sekundarnih disperzija. Npr. specifična haplogrupa M1, prisutna u Etiopiji i Somaliji, vjerovatno je rezultat povratnih migracija u istočnu Afriku.
Osim studija mtDNK varijacija i arheološke studije favorizuju concept ,,Route Southern Coastal,, kao glavni mehanizam primarnog naseljavanja Azije i širenju anatomskih modernih ljudi iz Afrke južnim koridorom u Aziju, a kasnije i u ostale dijelove svijeta (Armitage et al. 2011; Rosenberg et al 2011; Kivisild et al. 2003, 2004; Quintana-Murci et al. 1999 Stringer 2000). Da su obalna staništa medijum za probijanje ljudi iz istočne Afrike do Azije i Australije prvi put je predviđeno strane evolutivnog geografa Sauer-a 1962. godine, koji je smatrao da populacije koriste ovu rutu zbog prilagođenosti ekološkoj niži morskih obala.[5]
Nedavno objavljena filogenetička analiza (Fernandes et al. 2012) haplogrupa N1 i N2 (uključujući i reprezentativne haplogrupe I, W i X), koje izravno divergiraju iz ne-afričke izvorne makrohpalogrupe N, sugeriše da ove haplogrupe predstavljaju drevne relikte prvih humanih disperzija iz Afrike duž južne rute, ukazujući na njihovo drevno porijeklo u Arabijskom poluotoku, odakle su se najvjerovatnije širile prema Bliskom Istoku i Evropi prije 55-24.000 godina.
Druga ekspanzija se desila rutom prema sjevernoj Africi i Levant koridoru prije oko 50.000 godina (Bandelt et al. 2006; Torroni et al. 2006) i označena je kao sjeverna ruta ‘Northern Route’. Obuhvata disperzije kroz Bliski Istok, Arabiju do centralne Azije, odakle su se migracije dalje širile u pravcu Evrope, istočne i sjeveroistočne Azije (40-50.000), a nakon čega i u pravcu Amerike (do 15.000 godina) (sl.18). ,,Sjeverna ruta,, je obilježena ekspanzijom makrohaplogrupe N. Ovu rutu su obilježila tri glavna klastera, klaster kojeg čine W, I i N1B haplogrupe koje potiču direktno iz N1 i N2, odnosno, iz haplogrupe N (uočene u Evropi, B. Istoku i Kavkazu), zatim klaster rijetko prisutnih haplogrupa u Evropi X i A, te treći klaster kojeg čine: haplotipi B haplogrupe (karakterističan u istočnoj Aziji, Japanu i južnom Pacifiku), haplotipovi haplogrupa J i T, te haplogrupe H i V (treća skupina) čiji potomački derivati se mogu naći u Evropi, sjevrnoj Africi i centralnoj Aziji. I četvrti klaster U migrirao je u različite dijelove staroga svijeta sa najvišom frekvencijom svojih subhaplogrupa u Indiji (U2 i U7), sjevernoj Africi (U6 i U3) i Europi (U5).
Naseljavanje Evrope[uredi | uredi izvor]
Prema arheološkim podacima, prije oko 1,1 milion godina članovi roda Homo pojavili su se prvi put u Evropi. Prvo pojavljivanje recentnog čovjeka H.sapiens u Europi vezuje se za period od prije oko 45-50.000 godina, upravo nakon njegovih migracija iz područja Azije. Nakon ulaska u Evropu uslijedila je brza geografska disperzija ranih modernih ljudi. Jedan od zadataka molekularne antropologije je da procjeni kako je tekao genski tok prilikom naseljavanja Evrope i koji događaji iz prošlosti su imali dominatan efekat na genetičke varijacije recentnih evropskih populacija.
Komparacijom arheoloških podataka, obrazaca varijacija mtDNK i Y hromosoma (Oliveri et al. 2013; Higham et al. 2011; Soares et al. 2010; Barbujani & Goldstein 2004), prehistorija modernih ljudi u Evropi može se podijeliti u šest epizoda: 1) pionirske migracije iz Afrike prije oko 1 milion godina; 2) disperzije modernih ljudi iz Afrike, preko regiona Azije u ranom gornjem paleolitu; 3) ,,re-kolonizacije većine kontinenta iz glacijalnih refugijuma (pretežno južnih refugijuma) poslije ,,Last Glacial Maximum,,; 4) postglacijalna kolonizacija mezolitskim grupama (obilježavajući kraj Pleistocena i početak holocena); 5) neolitske disperzije sa B. Istoka; i 6) male skale migracija duž kontinenta uslovljene ekonomskim razmjenama u periodu željeznog doba. Svaka od ovih migracija je ostavila dio genetskog nasljeđa u evropskom genofondu modernih populacija.
Prema podacima studija evolucije mtDNK, dvije demografske ekspanzije imale su vjerovatno najveći uticaj struktuiranje današnjeg evropskog genofonda, to su kolonizacija prvim Homo sapiens sapiens u ranom gornjem paleolitiku (prije 45-50.000 godina) i neolitske ekspanzije iz B. Istoka (prije oko 10.000 godina). Između ove dvije ekspanzije, u posljednjem glacijalnom periodu (LGM), čiji je maksimum 18-20.000 godina, populacije su se povukle u nekoliko area tzv. ,,glacijalnih refugijuma,, iz kojih je nakon poboljšanja klimatskih uslova kroz period “Late Upper Paleolithic,” ili “Mesolithic,” nastupila re-kolonizacija evropskog kontinenta.
Pionirske migracije iz Afrike. Pionirske migracije vezuju se za period od prije oko 1 milion godina, kada su članovi roda Homo kročili prvi put na evropsko tlo (Carboneli et al. 2008). MtDNK (kao i Y hromosom) ukazuje na potpunu zamjenu arhaične hominidne loze anatomski modernim ljudima. Geografski, Evropa je smještena u zapadnom rtu Azije, izložena oscilacijama golfske struje, koje uzrokuju nagle klimatske fluktuacije, koje su vjerojatno imale uticaj na širenje i izumiranje arhaičnog H. sapinesa i njegovih populacija, uključujući i neandertalce. Disperzije modernih ljudi iz Azije. Prvi moderni ljudi (H.sapiens) prema paleoantropološkim podacima pojavljuju se u Europi u periodu ranog gornjeg paleolita (prije oko 40-45.000 godina) migrajući iz područja B. Istoka preko Levant koridora (Mellars, 2011, Higham et al. 2011). Analize mtDNK varijacija sugerišu na prvo naseljavanje Evrope anatomski modernim ljudima prije oko 50.000 godina. Najranije migracije iz B. Istoka prema Evropi obilježene su ekspanzijom haplogrupe U5 sa ancestralnim tipom sekvence HVSI-16270. Kao što je predhodno navedeno, haplogrupa U5 je najstarija evropska mtDNA haplogrupa koja se širila sa prvim evropskim naseljenicima u ranom gornjem paleolitu. Danas predstvalja samo oko 7% evropskog genskog bazena. Studija Malyarchuk et al. (2010) sugeriše da je prosječno koalescentno vrijeme za U5 25-30.000 godina, a nosioci U5 čija evolutivna dob 55-30.000 godina vezuju se za period najranijih migracija u Evropi. Haplogrupa ranog gornjeg paleolita pronađena je u velikoj frekvenciji (15-15%) kod Baski populacija, Skandinaviji i sjeverno-evropskim populacijama, dok u dosta manjoj frekvenciji (5-9%) u zapadnom Mediteranu, Alpama i sjevreocentralnoj i sjeverozapadnoj Evropi. Geografska distribucija haplogrupe U5 predstvalja relevantan pokazatelj prvobitnog naseljavanja sjevernih i zapadnih regiona Evrope. U modernim evropskim populacijama zabilježeno je prisustvo U5 subklastera manjeg koalescentnog vremena 11–13.000 godina (Tambets et al. 2003), sugerišući da su drevni U5 haplotipovi bili zajednički i za mezolitske i neolitske evropljane, posebno u centralnom i sjevernom dijelu Evrope.Druga najstarija haplogrupa u Evropi je U8 (oko 50.000 godina). Ostale komponente koje su širele u gornjem paleolitu iz pravca B. Istoka, su komponete srednjeg gornjeg paleolita (prije oko 26.000), to su haplogrupe HV*, U1, a posebno U2 i U4.
Re-ekspanzija iz glacijalnih refugijuma i postglacijalna kolonizacija. Nakon dolaska iz B. Istoka u Evropu humane populacije su se najviše koncentrisale na sjeveru i zapadu Evrope. Međutim, dramatične i brze klimatske promjene, čiji je vrhunac bio LGM (18-20.000), dovode do novih disperzija humanih populacija unutar evropskog kontinenta. Prema rezultatima studija mtDNK varijacija humane populacije kroz period LGM napuštaju sjeverne i centralne dijelove Evrope i koncentrišu se u ,,glacijalnim refugijumima,, pretežno u jugozapadnoj Evropi (France/Cantabria), zapadnoj mediteranskoj zoni i Ukrajini (planine centralne Rusije) (Periglacial zone), dok su ostali minorni refugijumi egzistirali između ovih (Dolukhanov 2000).
S obzirom da proces molekularne divergencije neizbježan, u glacijalnim refugijumima nastaju novi haplotipovi kao posljedice novih mutacija. Ovi haplotipovi, danas se označavaju glavnim signalima u modernom evropskom mtDNK bazenu re-ekspanzija iz glacijalnih utočišta. Novije studije sugerišu o ,,glavnim,, signalima postgalcijalne re-kolonizacije Evrope (prije oko 15.000 godina) i to, iz Franco-Cantabria refugijuma, kao što su H1, H3, H5, V i U5b1b (Pala et al. 2012), iz Ialijanskog poluostrva, haplogrupa U5b3 (Pala, et.al. 2009) i iz istočno evropskih planina U4 i U5a (Malyrchuk et al.2008; 2010). Ove mtDNK varijante, koje su se pojavile poslije ledenog doba sugerišu na moguće pravce postglacijalnih disperzija humanih populacija iz glacijalnih refugijuma prema sjevernoj, centralnoj i zapadnoj Evropi. Npr. rijetka haplohrupa u Evropi U5b3 (koalescentno vrijeme oko 10.000 godina) smatra se potencijalnim signalom re-kolonizcaije Evrope iz refugijuma u Italijanskom poluostrvu, čija se ekspanzija vezuje za Mediteransku obalu. Nekoliko mtDNK ,,lineage,, dominantnih u istočnoj Evropi, npr. subhaplogrupe U4 sugerišu na rezultate ekspanzija iz istočnih refugijuma, vjerovatno u Ukrajini. Haplogrupa U5 (podgrupe U5a i U5b) najviše su koncentrisane u populacijama na sjeveru (populacija Sammi, Finci i Estonci), a u nešto manjem stepenu u ostatku Evrope, te njihova distribucija i evolutivno vrijeme dobar su pokazatelj pravca početnih populacionih ekspanzija nakon LGM.
Sa pojavom deglacijacije evropskog kontinenta kroz ,,Latte Uper Paleolithic,, mtDNK haplogrupe širile su se različitim reginima Evrope. Osnovni doprinos tim kasnijim ekspanzijama u gornjem paleolitiku (prije oko 14.000 godina) dali su najveći subklasteri, kao što su haplogrupe H, K, T*, T2, W i X njihove subhaplogrupe. Ove haplogrupe su prevalente u zapadnom Mediteranu i Baski populacijama, sugerišući pravac migracija i repopulizacije jugozapad-sjever. Da je postglacijalna ekspanzija imala značajan uticaj u rekolonizaciji evropskog kontinenta svjedoče i rezultati novije studije Oliveri et al. (2013), koji ukazuju na tragove difuzije nekoliko subhaplogrupa I i W, prizašle iz perioda evropskog neolita, a koje su ograničene u Evropi.
Deglacijacija evropskog kontinenta započela je sa zagrijavnjem prije oko 15.000 godina, a stabilizovala se krajem 11.000 godina. U posljednjih oko 11.000 godina (Holocen) relativno stabilne klimatske prilike omogućile su mobilnost populacija i razmjenu genetskog materijala kada je došlo je do postepenog repopuliziranja Evrope iz svih glacijalnih refugijuma. U kontekstu rasprava o značaju pojedinih evropskih glacijalnih refugijuma za vrijeme LGM, molekularne disekcije i analize geografske distribucije zajedničkih evropskih haplogrupa H i U5 i autohtone evropske haplogrupe V, ukazuju da je postglacijalna ekspanzija paleolitskih populacija iz francuskog Cantabrian refugijumu (jugozapadna Evropa) mogla imati najznačajniji uticaj u repopulalizaciji evropskog kontinenta, posebno centralne i sjeverne Evrope, početkom Holocena.
Neolitske ekspanzije[uredi | uredi izvor]
Kao što klasični genetičari pripisuju uticaj neolitskih farmera u genskom toku iz Bliskog Istoka, također i studije mtDNK ukazuju na sudjelovanje neolitskih linija sa B. Istoka u mtDNK genskom toku Evropljana, ali u dosta manjem obimu. Molekularni podaci sugerišu da je kulturni i demografski uticaj sa Bliskog Istoka u neolitu bio veoma smanjen posebno na Iberijskom poluostrvu. Najznačajnji predstavnici neolitskih klastera, bile su mitohondrijalne haplogrupe J, T1, U3 , i nekoliko subhaplogrupa H i W. Najznačajnija genetička determinanta unesena prvim neolitskim farmerima u Evropu je N1a. Male zanemarljive imigracije u doba neolita (prije oko 9.000 i kasnije) čini se da su dovele mlađe sub-haplogrupe na evropsko tlo, kao što su J1b1, J2a i T1a, a i neke koje su veoma rijetke, kao što su R1, R2 i N1a. Podaci studije Pala et al. (2012) sugerišu da dvije zapadno-euroazijske haplogrupe J i T označene u predhodnim studijama kao markeri neolitskih ekspanzija iz B. Istoka, sa poboljšanjom rezolucijom ukuzuju da su se pojavile dosta ranije u Evropi, krajem Last Glacial Maximum.
Hipoteza o paleolitskom porijeklu Evropljana[uredi | uredi izvor]
Genski diverzitet u Evropi je interpretiran kao reflekcija različitih skala migracija, koje su se javljale kroz paleolitski (prije oko 45.000 godina), mesolitski (prije oko 17.000) i neolitski (prije oko 10.000) period. Genetički modeli prezentovani od mnogih populacionih genetičara i antropologa naglašavaju autohtono paleolitsko porijeklo savremenih evropljana. Ovaj koncept je zasnovan na pretpostavci da su današnji evropljani primarni potomci lovac-sakupljačkih grupa, koje su živjele u nekim regionima Evrope do prije oko 10.000 godina, tj. do perioda kada započinje naseljavanje neolitskim poljoprivrednim zajednicama. Prema ovom konceptu samo mali broj genskih lineage rezultat je neolitskih imigranata.
U inetrpretaciji rezultata istraživanja mtDNA linija (za evropske mtDNA linije i Y hromosom), na osnovu prostorne frekvencije sugerušu da evropski genski tok ima oko 80% paleolitskog i 20% neolitskog porijekla. Ove vrijednosti su dosta bliske proporciji opisanih varijacija na osnovu studije prvih klasičnih genetičkih markera.
Popularni model ,,demic difuzija,, (engl. demic-diffusion model) (Ammerman and Cavalli-Sforza 1964) sugeriše na jak uticaj neolitskih farmera B. Istoka na evropski genski bazen, te da velika proporcija današnjeg evropskog genskog bazena trebala bi biti izvedena iz neolitskih migranata. Ovaj model ukazuje na vrlo mali stepen miješanja migrantnih populacija iz B. Istoka sa mezolitskim evropskim naseljenicima i sporiju ekspanziju populacija iz izvornih neolitskih evropskih populacija. Suprotno ovom modelu, cultural-diffusion model sugerira transfer poljoprivredne tehnologije bez značajnih populacionih pomjeranja.
U odnosu na podatke klasičnih genskih lokusa, kao i drugih nuklearnih DNK markera, uključujući i varijacije Y hromosoma, početne studije o mtDNK varijacijama nisu podržavale demic-difuzija model. Nekoliko najznačajnijih studija koje su se bazirale na procjenu porijekla evropskih mtDNK varijacija sugerirale su da većina evropskih mtDNK sekvecni ima porijeklo iz gornjeg paleolita, da su unesene u Evropu kroz nekoliko valova u gornjem paleolitu, da je bio prisutan bottleneck efekat ili efekat osnivača u acocijaciji sa LGM prije oko 20.000 koji je doveo do postglacijalne divergencije velikog broja odijeljenih mitohondrijskih linija i da je neolitska komponenta zastupljena manje od jedne četvrtine u mtDNK toku evropskih populacija. Većina postojećih evropskih mtDNK varijacija ima porijeklo posljednje glacijalne ekspanzije (prije oko 14.000 godina) koja se desila unutar Evrope, sa oko 10% udjela mtDNK varijacija iz perioda ranog gornjeg paleolita. Kao što je već predhodno napomenuto, paleolitske migracije iz jugozapadne Evrope prema ostatku Evrope imale su najveći udio u mitohondrijalnom genskom toku evropskih populacija. Nivo diverziteta i geografska distribucija autohtonih mtDNK evropskih linija otkrivaju da se paleolitska populaciona ekspanzija iz jugozapada prema centralnoj i sjevernoj Evropi prije oko 10-15.000 godina vjerovatno imala glavni uticaj na rekolonizaciju kontinenta.
Nakon popularne teorije modela demografska difuzija raspravljalo se o relativnom doprinosu neolitskog genskog bazena u ukupnom evropskom mtDNK genofondu. Jedna od studija koja je nastojala odgovoriti na krucijalno pitanje, da li su evropljani potomci prvih evropskih farmera iz neolitskog perioda od prije 7.500 godina ili paleolitski lovci-sakupljači koji su bili prisutni u Evropi prije 40-50.000 godina, bila je analiza drevnih mtDNK iz 24 neolitska fosilna ostataka lokacija u Njemačkoj, Austriji i Mađarskoj (Haak 2005). Procjenjena starost pronađenih uzoraka je od 7500 do 7000 godina, što odgovara Linear Pottery Culture (Linearbandkeramik ili "LBK") (keramička kultura) koja se vezujue za početak ranih poljoprivrednih kultura u centralnoj Evropi. Od 24 uzorka koliko je analizirano u pomenutoj studiji jedna trećina je pripadala haplogrupama čije se rasprostranjenje vezuje za Evropu, B. Istok i centralnu Aziju (H, V, T, K, J i U3). Zbog nedostatka temporalne ili gografske distribucije ove haplogrupe nisu bile dovoljno informativne za procjenu paleolitskog ili neolitskog pitanja porijekla. Stoga je fokus istraživača ove studije bio šest uzorka pripadajuće haplogrupe N1a, linije haplogrupe N, koja je danas rijetko zastupljena među evrpljanima. N1a je pronađena u visokoj frekvenciji (8%-42%) i to pretežno u lokacijama u Mađarskoj i Njemačkoj, dosta većoj nego među današnjim Evropljanima.
Na osnovu dobijenih podataka, autori su zaključili da je rana neolitska agrikultura imala ograničen doprinos u genskom toku materalnih linija modernih evropljana. Prema ovim podacima, moderni evropljani se ne bi mogli smatrati potomcima prvih evropskih faremara, nego direktni potomci autohtonih paleolitskih lovac-sakupljača. Rezultati ove studije ukazuju na to da je haplogrupa N1a bila prisutna u Europi prije 7000 godina i to u ranoj LBK kulturi na ograničenom području u sredini Podunavlja, međutim doživjela je nestanak za gotovo dvije hiljade godina nakon pojave poljoprivrede. Mogući faktori koji su mogli uticati na nestanak linije N1a iz modernih evropskih loza su genetički drift i postneolitske ekspanzije. Noviji podaci sugerišu na naknadna populaciona kretanja iz B. Istoka prije oko 5000 godina, koja su mogla imati uticaj na paleolitske popualcije i da je kasnoneolitička kultura mogla imati ključnu ulogu u oblikovanju moderne srednjoeuropske genetske raznolikosti. Paleolitsko porijeklo evropljana se relativno dugo zadržalo i tokom kasnijeg mitohondrijalnog genskog toka, bez obzira na velika demografska pomjeranja farmera iz B. Istoka kroz neolitsku eru. Još uvijek nije precizno jasno u kojoj mjeri su prve migracije u Evropi oblikovale mtDNK bazen modernih evropljana, iako su kasnije imigracije iz B. Istoka mogle bitno uticati na genetičku strukturu evropskih populacija. Nekoliko skala migracija porijeklom sa Bliskog istoka od perioda ranog gornjeg paleolita pa do neolita, te i u novije vrijeme vjerojatno su doveli u Europu veliki broj nezavisnih osnivačkih loza, inkorporirani unutar osnovnih haplogrupa pre-HV, JT, U, N1, N2, i X. Novije studije ukazuju na neočekivani i značajan genski diskunuitet koji postoji između savremenih evropljana i njihovih paleolitskih predhodnika. Prema tome, savremene evropljane ne bih trebalo posmatrati u potpunosti odvojene od ranih neolitskih popljoprivrednika ili autohtonih paleolitskih stanovnika Evrope. Mitohondrijalni genski bazen je vrlo sličan širom evropskog kontineta i ne pokazuje značajno odstupanje čak i područjima gdje žive izolavne populacije kao što su Baski. Genetička struktura Evropljana iz perspektive mtDNK rezultirala je iz različitih prahistorijskih populacija i njihovih kultura koje su živjele na ovim prostorima u različitim vremenskim razdobljima.
Trebalo bi napomenuti jednu značajnu opservaciju. Model evropskog genetičkog porijekla koristi genetičku rekonstrukciju većinom zasnovanu na podacima savremenim populacijama i često ne promoviše studije drevnih DNA. Najpouzdanije analize uključuju korištenje ne samo uzoraka mtDNK savremenih populacija nego i drevnih mtDNK u rekonstrukciji genetičke historije stanovništva.
mtDNK neandertalaca[uredi | uredi izvor]
Većina hominidnih fosila su još uvijek izvan dometa istraživačkih mogućnosti molekulske evolucije, upravo zbog degradacije uzoraka i nedovoljne količine DNK. Od momenta kada su pronađeni prvi fosilni ostaci neanertalaca, genetičke relacije između neandertalaca i modernih ljudi su predmet mnogih debata. Neandertalci su u periodu 150.000-30.000 godina naseljavali gotovu cijelu Evropu, dio Azije i područja Srednjeg Istoka. Prema literaturnim podacima, moderni ljudi i neandertalci su zajedno egzistirali u Evropi prije 30-40.000 godina. Prema ovom saznanju, puno je kontraverzi oko nestanka nenadertlaca i mogućeg miješanja sa modernim ljudima. Studije koje su sprovedene na mtDNK izolovanoj iz drevnih fosilnih ostataka neandertalaca, omogućile su direktan uvid u mitohondrijalnu raznolikost i identifikaciju haplotipova koji su se uspjeli sačuvati do danas. Deset godina poslije prvog publikovanog rada Cann et al. (1987) koji podržava model out—Africa objavljena je studija pod nazivom “Neanderthal DNA sequences and the origin of modern humans” (, u kojoj su date prve procjene o genetskoj udaljenosti između nendertalaca i modernih ljudi. Studije izvedene na mtDNK fosilnih kostiju otkrivenih u Njemačkoj 1856. i Hrvatskoj, sugerirale su da je mtDNK sekvenca neandertalaca izvan ranga diverziteta humane DNK, što ukazuje da neandertalci nisu bitno uticali na današnji humani mitohondrijalni genski bazen. Također, studija Ovchinnikov et al. (2000) analizira diverzitet fosilnog ostatka iz sjevernog Kavkaza starog oko 29.000 godina i otkriva da sekvence 3,48% odstupa od sekvence fosilnog ostatka pronađenog u Njemačkoj. Također, i ova studija podržava koncept da je neandertalska mtDNK evoluirala odvojeno od modernog čovjeka, i da se evolucijsko odvajanje nendertalaca od modernih ljudi desilo prije oko najmanje 500.000 godina.[7]
Konstatacija o genetskoj udaljenosti između neandertalaca i modernih ljudi ne pruža dokaz za multiregionalnu hipotezu humane evolucije, ukazujući na to da su anatomski moderni ljudi i neandertalci bili različiti. Međutim, ne treba izbjeći ni pretpostavku eventualnog miješanja ranih modernih ljudi i neandertalaca, ali zbog značajne uloge genetičkog drfita ili genetičkog uskog grla ovi haplotipovi vjerovatno nisu uspjeli preživjeti do danas. Ono što relativno značajno otežava detaljniju analizu je kontaminacija fosilnih ostataka i nedovoljna količina uzorkovanog materijla ranih modernih ljudi, što bitno utiče na donošenje nekih preciznijih sudova o ulozi mitohondrijalne sekvence neanderlataca u ukupnom genofondu modernog čovjeka. Dalji izvještaji poređenja mtDNK neandertalaca i modernih ljudi vjerovatno će dovesti do preciznijeg zaključka.
Reference[uredi | uredi izvor]
- ^ Oven M., Kayser M. (2009): Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation. Human Mutation, 30 (2): E386–94.|pmid=18853457 |doi=10.1002/humu.20921
- ^ [http: / /mbe.oxfordjournals.org/content/21/11/2012.long "Disuniting Ujednačavanje: A Pied kladističkih Canvas mtDNK haplogrupa H u Evroaziji"]
- ^ Disuniting Uniformity: A Pied Cladistic Canvas of mtDNA haplogroup H in Eurasia
- ^ a b c d e Ahmić A., Silajdžić E. (2014): Mitohondrijalna DNK i Y hromosom u filogenetičkim studijama humanih populacija. OFF-SET, Tuzla, ISBN 978-9958-31-184-0.
- ^ a b http://www.mitomap.com.
- ^ Brook, Kevin Alan (2022). The Maternal Genetic Lineages of Ashkenazic Jews (jezik: engleski). Academic Studies Press. str. 15. doi:10.2307/j.ctv33mgbcn. ISBN 978-1644699843.
- ^ Ahmić A., Silajdžić E. (2014): Mitohondrijalna DNK i Y hromosom u filogenetičkim studijama humanih populacija. OFF-SET, Tuzla, isbn 978-9958-31-184-0.
Također pogledajte[uredi | uredi izvor]
Vanjski linkovi[uredi | uredi izvor]
- Mitohondrijsko filogenetičko stablo
- Ian Logan's Mitochondrial DNA Site
- Mannis van Oven's PhyloTree.org
- PhyloD3 - D3.js-based phylogenetic tree based on PhyloTree Arhivirano 29. 10. 2020. na Wayback Machine
- Mitochondrial haplogroup skeleton
- Vincent Macaulay's Mitochondrial haplogroup motifs
- Cambridge DNA's An mtDNA view of the peopling of the world by Homo sapiens
- John S. Walden's Map of macro-haplogroup N
- List of mtDNA haplogroup projects
- MitoTool: a web server for the analysis and retrieval of human mitochondrial DNA sequence variations Arhivirano 19. 6. 2016. na Wayback Machine
- HaploGrep: mtDNA haplogroup determination based on PhyloTree.org Arhivirano 12. 6. 2016. na Wayback Machine
- HaploFind - fast automatic haplogroup assignment pipeline for human mitochondrial DNA Arhivirano 11. 6. 2016. na Wayback Machine
{kind=link}
{kind=link}