
CCDC142
| CCDC142 | |||||||
|---|---|---|---|---|---|---|---|
| Identifikatori | |||||||
| Aliasi | |||||||
| Vanjski ID-jevi | GeneCards: [1] | ||||||
| Ortolozi | |||||||
| Vrste | Čovjek | Miš | |||||
| Entrez |
|
| |||||
| Ensembl |
|
| |||||
| UniProt |
| ||||||
| RefSeq (mRNK) |
|
| |||||
| RefSeq (bjelančevina) |
|
| |||||
| Lokacija (UCSC) | n/a | n/a | |||||
| PubMed pretraga | n/a | n/a | |||||
| Wikipodaci | |||||||
| |||||||
Protein upredene zavojnice 142 (CCDC142) je gen koji kod ljudi kodira protein CCDC142. Gen CCDC142 se nalazi na hromosomu 2 (na 2p13), obuhvata 4.339 parova baza i sadrži devet egzona. Gen kodira domen upredene zavojnice 142 (CCDC142), čija funkcija još nije dobro shvaćena.[1][2] Postoje dvije poznate izoforme CCDC142.[1] CCDC142 proteini proizvedeni iz ovih transkripata imaju veličinu od 743 do 665 aminokiselina i sadrže signale koji ukazuju na kretanje proteina između citosola i jedra.[1] Iako funkcija ovog proteina nije dobro shvaćena, zna se da sadrži domen upredene zavojnice i RINT1 TIP1 motiv koji se nalazi unutar domena upredene zavojnice.[3][4]
Lokus[uredi | uredi izvor]

CCDC142 se nalazi na 2p13.1 lancu hromosoma 2, sa genomskom sekvencom koja obuhvata baze od 74,472,832 do 74,483,230.[1] Kodirajuća regija duga je 8.292 bazna para, koja kodira dvije izoforme proteina dužine od 743 do 665 aminokiselina.]]|[1] Na telomernoj strani, CCDC142 praćen je genima MOGS i MRPL53, a na centromernoj, slijede geni C31, LBX2, LBX2-AS1 i PCGF1.[1]
iRNK[uredi | uredi izvor]
U Homo sapiens, gen CCDC142 kodira dvije alternativno prerađene izoforme iRNK, zvane izoforma 1 i izoforma 2.[3] Obje ove izoforme imaju 9 egzona. Izoforma 1 je duža od ta dva, duga je 4339bp, dok je izoforma 2 duga 2253bp.[3] Obje izoforme imaju po devet egzona. Izoforma 1 je duža i ima 4.339 bp, dok je izoforma 2 duga 2253 bp.[3] Glavna razlika između dviju izoformi je što izoforma 2 ima kraći egzon 9 and 3' UTR.[3] Izoforma 1 je duža varijanta gena i kodiranog proteina, na koje se ovaj članak i odnosi.[1]
Protein[uredi | uredi izvor]

Primarna struktura, varijante transkripta i izoforme[uredi | uredi izvor]
Glavna izoforma proteina CCDC142 duga je 743 aminokiseline, a druga 665 aminokiselina. Razliku u dužini u potpunosti čine aminokiseline koje nedostaju na C-terminalu izoforme 2.[1]
Domeni i motivi[uredi | uredi izvor]
Predviđeni domen upredene zavojnice CCDC142 je od aminokiselina 308–719.[2] RINT1 TIP1 motiv je također prisutan od aminokiselina 490–621 . RINT1 TIP1 je porodica koja uključuje RINT-1 (protein uključen u kontrolu kontrolnih tačaka izazvanih radijacijom) i TIP-1 (protein kvasca koji je uključen u Golgijev transport).[4] Dodatnih ~250 aminokiselina koje se nalaze u udaljenom ortologu CCDC142 proteina ne nalazi se u ljudskom genomu blizu gena CCDC142.
Posttranslacijske modifikacije[uredi | uredi izvor]
Predviđeno je da CCDC142 ima šest mjesta fosforilacija, četiri mjesta metilacija , jedno mjesto palmitoilacija, [jedno [SUMO protein|sumoilacija]] i jeda slabi jedarno lokalizacijski signal.[5][6][7][8][9] Ove modifikacije ukazuju da je CCDC142 lokalizovan u nukleusu i citosolu.
Predviđanje strukture[uredi | uredi izvor]
Sekundarna struktura CCDC142 sadrži samo α-heliks kako je predviđeno programima Quick2D i Phyre2 .[10][11] Predviđeno je da CCDC142 sadrži osam konzerviranih α-heliksa, od kojih se šest nalazi u području zavojnice proteina.[10][11] Predviđena tercijarna struktura CCDC142 sadrži veliki zavojni domen od aminokiselina 308–719.[2][12]

Ekspresija[uredi | uredi izvor]
Promotori i regulatorni faktori[uredi | uredi izvor]
Region promotora za CCDC142 identifikovan je korištenjem programa El Dorado na Genomatixu, obuhvata baze 74482896–74483908 u hromosomu 2.[13] Ova regija od 1.013 bp proteže se uzvodno 1071–58 bp od startnog kodona CCDC142.[13] Postoji regija u promotoru koja vezuje veliki broj Krueppelolike transkripcijske faktore i BED-proteina cinkovog prsta.[13] Ova regija nema jednonukleotidnih polimorfizama (SNP).[14] Mnogi transkripcijski faktori koji se vezuju za promotorsku regiju CCDC142 imaju funkcije u supresiji tumora, neurogenezi, oštećenju DNK i fotorecepciji.[13] Ova promotorska regija također ima ulogu u radu LTR-TATA kutije sisarskog C-tipa koji se preklapa sa početnom lokacijom transkripcije gena.[13]
RNK vezujući proteini[uredi | uredi izvor]
Brojni su mogući proteini koji se vezuju za RNK; vezuju se i za 3' UTR i neprevedene regije iRNK CCDC142. Mjesta vezanja za proteine PABPC1 i RBMX javljaju se na visokoj frekvenciji u 3’ UTR, sa 49. odnosno 21. mjestom.[15]
Ekspresija[uredi | uredi izvor]
- Allen Human Brain Atlas Expression of CCDC142
-
 Bočni izgled
Bočni izgled
Crveno=Niska ekspresija11 -
 Frontalni izgled
Frontalni izgled
Zeleno=Visoka ekspresija11


Iznad su podaci o ekspresiji Alenovog atlasa ljudskog mozga, ekspresiji na CCDC142, pri čemu crvena označava nižu ekspresiju, a zelena označava veću.[16] U ljudskom mozgu otkriveno je da je CCDC142 slabo eksprimiran u cerebelumskoj kori, talamusu i hipotalamusu. CCDC142 je takođe visoko izražen u substantia nigra, ponsu, klaustrumu i mezencefalonu.[16] Također postoji relativno viši nivo ekspresija CCDC142 u ustima i timusu.[17]
- NCBI GEO Expression Data
-
 MEKK 2/3 eksperiment nokauta 13
MEKK 2/3 eksperiment nokauta 13 -
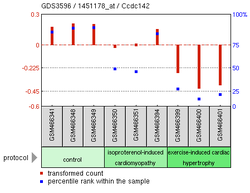 Eksperiment oštećenja miokarda13
Eksperiment oštećenja miokarda13 -
 Eksperiment nadekspresije SNAI13
Eksperiment nadekspresije SNAI13



Gore navedeni eksperimentalni podaci pokazuju mnoge moguće nalaze za CCDC142.[18] Prekomjerna ekspresija SNAI1, proteina cinkovog prsta, povezana je sa smanjenjem ekspresije u ljudskom CCDC142.[19] Mišji nokaut-gen MEKK 2/3, koji pomaže u regulaciji diferencijacije T-pomoćnih ćelija, također je pokazao smanjenu ekspresiju CCDC142.[20] Još jedan eksperiment na Mus musculus fokusiran na kardiomiopatiju kod miševa pokazao je niže nivoe CCDC142 kod miševa sa oštećenim ćelijama miokarda.[19]
Funkcija i biohemija[uredi | uredi izvor]
Kompozicija[uredi | uredi izvor]
CCDC142 ima relativno tipsku raspodjelu aminokiselina, u poređenju sa drugim proteinima Homo sapiens.[21] Međutim, neke varijacije su zabeležene među ortolozima.[21] Leucin je prisutan u velikim količinama u odnosu na druge proteine (na preko 15% proteina), a asparagin u malim količinama u odnosu na druge proteine (sa manje od 0,7% proteina).[21]
Upredenozavojnički domen i motiv RINT1_TP1 CCDC142 sadrže veće količine leucina u odnosu na ostatak proteina (na preko 16,6% regije), veće količine glutamina (na preko 8,4% regije) i slično niske količine asparagina (na manje od 0,7% regije).[21]
Interaktivni proteini[uredi | uredi izvor]
Nisu pronađene interakcije proteina sa CCDC142.
Konzervacija[uredi | uredi izvor]
Paralozi[uredi | uredi izvor]
CCDC142 nema paraloga u Homo sapiens.
Ortolozi[uredi | uredi izvor]
Ispod je tabela raznih ortologa CCDC142 čiji je identitet proteinske sekvence upoređen sa sekvencom aminokiselina proteina Homo sapiens. CCDC142 ima više od 73% sličnosti aminokiselina kod ostalih sisara, ali je manje konzerviran kod drugih kičmenjaka i beskičmenjaka.[21]
| Rod i vrsta | Uobičajeno ime | Datiranje divergencije od ljudske loze (milioni godina) | Identitet (%) |
| Homo sapiens | Čovjek | 0 | 100 |
| Pan troglodytes | Obični čimpanza | 6,6 | 96 |
| Gorilla gorilla gorilla | Gorila | 8,9 | 98 |
| Jaculus jaculus | Mali egipatski skočimiš | 90,9 | 73 |
| Bos mutus | Jak | 97,5 | 74 |
| Eptesicus fuscus | Veliki smeđi šišmiš | 97,5 | 74 |
| Python bivittatus | Mjanmarski piton | 320,5 | 36 |
| Gallus gallus | Kokoš | 320,5 | 35 |
| Haliaeetus leucocephalus | Ćelavi orao | 320,5 | 33 |
| Anolis carolinensis | Gušter karolinska anola | 320,5 | 33 |
| Calidris pugnax | Ptica ruf | 320,5 | 32 |
| Xenopus tropicalis | Zapadna kandžasta žaba | 355,7 | 33 |
| Callorhinchus milii | Australijska duh-ajkula | 429,6 | 36 |
| Lepisosteus oculatus | Pjegava riba gar | 429,6 | 34 |
| Esox lucius | Sjeverna štuka | 429,6 | 33 |
| Danio rerio | Zebrica | 429,6 | 33 |
| Lingula anatina | Repasta dagnja | 847 | 29 |
| Crassostrea gigas | Pacifička ostriga | 847 | 29 |
| Octopus bimaculoides | Kalifornijski dvopjegi oktopus | 847 | 27 |
| Drosophila melanogaster | Vinska mušica | 847 | 23 |
Filogenija[uredi | uredi izvor]
CCDC142 je blisko srodan kod sisara, mehkušaca, vodozemaca, gmizavaca i ptica i riba.[21] Gen CCDC142 datira još od Drosophila melanogaster, koji se odvojio od ljudske loze prije 847 miliona godina. CCDC142 je mutirao većom brzinom i od citohroma C (visoko konzervirani protein) i fibrinogena A (protein koji brzo mutira). Ovo ukazuje da je CCDC142 gen koji brzo mutira sa sve većom stopom mutacije (odnosno, evolucije) tokom vremena.

Klinički značaj[uredi | uredi izvor]
Patologija i bolesti[uredi | uredi izvor]
Povećanje broja kopija u lokusima CCDC142, uključujući 25 drugih gena, eksprimiralo je fenotip kašnjenja u razvoju i značajne razvojne ili morfološke promjene.[22] Jedan rezultat sa gubitkom broja kopija u lokusima CCDC142, uključujući 29 drugih gena, pokazao je fenotipove nizak rast, abnormalni oblik lica, odgođeni razvoj govora i jezika, preklapanje prstiju, unutarmaternično ograničenje rasta, otvoreni ductus arteriosus i odložen razvoj grube motorike.[22] Međutim, efekt CCDC142 je možda bio zbunjujući za ove fenotipove jer su postojale i abnormalnosti u mnogim drugim dijelovima genoma.
Mutacije[uredi | uredi izvor]
Postoji veliki broj jednonukleotidnih polimorfizama koji se nalaze u genu CCDC142. Neki od njih u regiji promotora i 5' UTR-a su unutar sidrenih sekvenci za transkripcijski faktor i utiču na vezivanje transkripcionog faktora ako se promijene.
Postoji mnogo SNP-ova u kodirajućoj sekvenci proteina koji mijenjaju aminokiselinski sastav CCDC142. Jedan od SNP-ova sa visokom stopom prevalencije u populaciji (1,8%) je značajan po hemijskim promjenama, sa tirozinskim pomakom u asparagin na 548. aminokiselini.[14]
Postoje i brojni SNP-ovi koji se nalaze u velikoj neprevedenoj 3' UTR gena, pri čemu se mnogi od njih vezuju za područja koja sadrže strukture drška petlja u iRNK. SNP sa stopom prevalencije od 7,7% (guanin do adenozina na bp 4285.) je u 3' UTR-u, ali nije lociran u konzerviranoj regiji petlja-drška.[14]
Ovi SNP-ovi su označeni u konceptnom prevodu koji se nalazi u odeljku protein iznad.
[uredi | uredi izvor]
- Distant Ortholog Multiple Sequence Alignment of CCDC142
-

-

-
 Ljubičasto=Hemijski slične aminokiseline
Ljubičasto=Hemijski slične aminokiseline
Plavo= Iste aminokiseline -
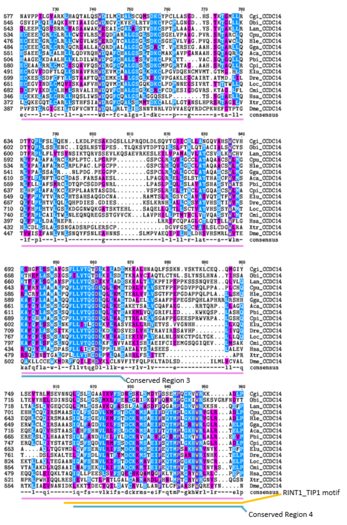
-






U višestrukom poravnavanju sekvenci iznad (kreiranom pomoću programa CLUSTALW i TEXSHADE na SDSC Biology Workbench), organizmi su označeni prvim slovom njihovog roda i prva dva slova njihove vrste. Cijeli protein CCDC142 je visoko konzerviran kod sisara.[21] Regije koje sadrže ljudski domen upredene zavojnice i region motiva RINT1_TIP1 su visoko konzervirane u udaljenim homolozima.[21] Čak 12 od 15 aminokiselina koje se poklapaju sa svim organizmima u ovoj regiji su nepolarne.[21] Konzervirano područje 1 sadrži uglavnom nepolarne aminokiseline,[21] područje 2 sadrži uglavnom nepolarne i bazne aminokiseline. Konzervirana regija 3 sadrži i polarne i nepolarne aminokiseline.[21] Konzervirana regija 5 sadrži uglavnom nepolarne i bazne aminokiseline.[21]
Dodatne informacije o transkripcijskom faktoru[uredi | uredi izvor]
Reference[uredi | uredi izvor]
- ^ a b c d e f g h i "CCDC142 coiled-coil domain containing 142 [Homo sapiens (human)] – Gene – NCBI". www.ncbi.nlm.nih.gov. Pristupljeno 1. 5. 2016.
- ^ a b c "coiled-coil domain-containing protein 142 [Homo sapiens] – Protein – NCBI". www.ncbi.nlm.nih.gov. Pristupljeno 1. 5. 2016.
- ^ a b c d e "CCDC142 – Coiled-coil domain-containing protein 142 – Homo sapiens (Human) – CCDC142 gene & protein". www.uniprot.org. Pristupljeno 1. 5. 2016.
- ^ a b "SSDB Motif Search Result: hsa:84865". www.kegg.jp. Pristupljeno 1. 5. 2016.
- ^ "NetPhos 2.0 Server". www.cbs.dtu.dk. Pristupljeno 1. 5. 2016.
- ^ "Memo:Protein Methylation Prediction". www.bioinfo.tsinghua.edu.cn. Arhivirano s originala, 14. 3. 2016. Pristupljeno 1. 5. 2016.
- ^ ":::NBA-Palm – Prediction of Palmitoylation Site Implemented In Naive Bayesian Algorithm:::". www.bioinfo.tsinghua.edu.cn. Arhivirano s originala, 9. 6. 2016. Pristupljeno 1. 5. 2016.
- ^ "SUMOplot™ Analysis Program | Abgent". www.abgent.com. Pristupljeno 1. 5. 2016.
- ^ "NLS_Mapper". nls-mapper.iab.keio.ac.jp. Arhivirano s originala, 22. 11. 2021. Pristupljeno 1. 5. 2016.
- ^ a b Kelley, Lawrence. "PHYRE2 Protein Fold Recognition Server". www.sbg.bio.ic.ac.uk. Pristupljeno 1. 5. 2016.
- ^ a b Remmert, Michael. "Quick2D". toolkit.tuebingen.mpg.de. Pristupljeno 1. 5. 2016.
- ^ a b c "I-TASSER server for protein structure and function prediction". zhanglab.ccmb.med.umich.edu. Pristupljeno 1. 5. 2016.
- ^ a b c d e "Genomatix – NGS Data Analysis & Personalized Medicine". www.genomatix.de. Arhivirano s originala, 24. 2. 2001. Pristupljeno 1. 5. 2016.
- ^ a b c snpdev. "SNP linked to Gene (geneID:84865) Via Contig Annotation". www.ncbi.nlm.nih.gov. Pristupljeno 1. 5. 2016.
- ^ "RBPDB: The database of RNA-binding specificities". rbpdb.ccbr.utoronto.ca. Pristupljeno 1. 5. 2016.
- ^ a b "Microarray Data :: Allen Brain Atlas: Human Brain". human.brain-map.org. Pristupljeno 1. 5. 2016.
- ^ "EST Profile – Hs.430199". www.ncbi.nlm.nih.gov. Pristupljeno 1. 5. 2016.
- ^ geo. "Home – GEO – NCBI". www.ncbi.nlm.nih.gov. Pristupljeno 1. 5. 2016.
- ^ a b "GDS3596 / 1451178_at". www.ncbi.nlm.nih.gov. Pristupljeno 1. 5. 2016.
- ^ "GDS4795 / ILMN_3023885". www.ncbi.nlm.nih.gov. Pristupljeno 1. 5. 2016.
- ^ a b c d e f g h i j k l "SDSC Biology Workbench".
- ^ a b ClinVar. "No items found – ClinVar – NCBI". www.ncbi.nlm.nih.gov. Pristupljeno 5. 5. 2016.